前几天看到“以科学方法辩论”发的一个贴子《生物学历史上最大的骗局》,贴中有这样的一段:
“这里提出一个宽松的公式,根据突变机率计算进化产生新物种的概率:
P =(M ? C ? L ? B ? S)^N――式1-1……”
对于生物学我没有太多研究,贴子里各位说的也都不少了。不过楼主提到的这个概率问题我倒可以发表些看法。
文中的公式中没有对各参数项给出明确解释,所幸这个公式还有其他的版本,如下:
“P=(M * C * R * E * S)^n (注: n 代表 n次幂) ――式1-2
通俗地说,就是一个物种的某个体发生了突变(机率M=0.001),并且突变后的基因与自身其它基因在不同层次的产物上可以相容(宽松估计C=0.01),而且在生存竞争中该个体能够存活,有繁殖的机会(R=0.1),而且突变恰好有纵向进化的意义(这种情况至今没有发现,某些进化论者估计为E=0.001),而且突变基因在种群中得以扩大的,(宽松的估计为S=0.1);因为新物种的形成需要一系列新基因的出现,假设要10个(幂指数n=10,实际物种间绝对没有这么小的基因差异)那么进化出一个新物种的概率
P=(0.001×0.01×0.1×0.001×0.1)^10 = 10^(-100) (-100次幂)
按照一年繁殖10代,种群个体数为1000,相应的进化所需要的时间极为宽松的计算也需要‘10的96次幂’年。目前科学认为宇宙中所有基本粒子总数只有10的70次方个,宇宙年龄不会超过200亿(2乘10的10次幂年),进化一个新物种的时间,是宇宙的年龄的自乘10亿次,足见进化是绝不可能的。……”
这个公式在很多地方都被引用过,目的都是说明进化不可能,而反对进化论,造成了不少认识混乱。所以一反以前先论证后结论的做法,这次我们开始就指出,上面那个概率式及其应用都是错误的,从而其推论也就是完全错误的。
公式中给出的各项的值从何而来、是否准确姑且不论了,就算都是对的吧。我们主要看公式本身和应用方法是否正确。首先,多余项,项S(突变基因在种群中得以扩大)。从公式及前后文来看,应该是只计算一代繁殖的概率,且只考虑子代单一个体,所以项S是多余的项。贴中的这个版本修正了这一项,“突变基因在种群中不被丢失、稳定、扩大,一旦丢失就又得重来(宽松的估计为10e-2)”,看起来好象说通了,实际上不通,因为刚才说过了,没有考虑再下一代,所以也就没必要考虑“丢失”。因此式1-2应该精简为:
P=(M*C*R*E)^n――式1-3
还有的项根本无意义,重复列项。如“突变恰好有纵向进化的意义”(项E),大家应该知道进化是没有方向性的,唯一使基因突变保留下来,外显为物种进化的决定因素就是自然选择,而式1-3中的项R和项C就是为保证适应自然的选择而设置的。所以项E可以去掉。式1-3可精简为:
P=(M*C*R)^n――式1-4
其次,存在概念模糊,时而指物种,时而指基因。如该式中M给出时,设定为物种个体突变的概率(已包含了相应的基因变异的概率),后面的n又设定为基因个数,前后概念混乱,导致基因变异的概率被两次引用,大大降低了事件发生的概率。要纠正这个错误,就要统一前后概念。对于文中公式来说,要么M项的定义是正确的,那就要去掉后面的n,就是说上式的结果不是10^(-100),而是10^(-6);要么M项的定义错误,应该定义为“某个基因发生突变的概率”。按前者结论远没文中吹嘘的那么惊人,那我们姑且认为后者是正确的,这样式1-4中括号内的部分,就是亲代生物的一个基因发生可遗传的变异的概率。而生物不可能只有一个基因,因此需要考虑所有基因中发生变异的概率。设该生物的遗传物质包括m个基因,则单一个体子代发生一个可遗传的基因突变的概率,才是明晰的、可以拿来作为计算新物种出现概率的依据。我们记为P1:
P1=C(1,m)*(M*C*R)*(1-M*C*R)^(m-1)――式2-1
注:式中C(1,m)为组合数。C(a,b)=b*(b-1)*(b-2)*……*(b-a+1)/a!
因为M*C*R很小,而m很大,上式可简化为
单一个体子代发生一个可遗传的基因突变的概率P1=m*(M*C*R)――式2-2
然后我们看看式1-4,就是P=P1^n是什么意思。从文中看,是因为“新物种的形成需要一系列新基因的出现”给出了n,用来表示新旧物种基因差别。而前面我们一直在讨论一代繁殖中单一个体的变异,这意味着什么?这意味着在一代繁殖中就出现新的物种!公式提出者给出的条件果然是太“宽松”了,你想啊,要让猫直接生出狗来,所需要的基因突变何止10个!大家看出荒谬来了吧。进化不是一下就完成的,需要一个长期的过程,物种变化所需的变异也决不是一次同时出现的,是个长期积累的过程。因而该项(n)的设置及使用根本就是错误的。要考察新物种出现的概率,就要综合考虑种群多代繁殖中基因突变的固定和组合,直到足够出现新物种。
自然界中生物的繁殖方式大体可分为两类,无性生殖和有性生殖,其后代基因的分配和组合固定的方式是不同的,因此应该分两种情况分析。我们首先看无性生殖。我们用无性生殖中最有代表性的分裂生殖来进行研究,一个细胞可分裂为两个(1代),再次分裂为四个(2代)……在经i代后其后代总数为2^i个,因为只考虑了每次出现一个新基因的情况,所以应有i>=n。如果每次出现不止一个新基因,其进化的过程应该更快,我们这里只考虑最坏的情况。计算时不再考虑经过生存竞争的减少,因为式2-2中的项R=0.1已经考虑进去了。因而在i代时,单一个体产生满足至少n个基因差异的概率为
P2=∑C(k,i)*(P1^k)*[(1-P1)^(i-k)] (式中k=n,n+1,n+2,……,i)――式3-1
此式计算复杂,可适当简化,如设代数i>=100(对于分裂生殖的生物来说,这并不需要多长时间),可采用泊松定理,则有单一个体产生满足至少n个基因差异的概率为:
P2=∑{(λ^k)*[e^(-λ)]/k!} (式中k=n,n+1,n+2,……,i;λ=i*P1)――式3-2
此式的值可直接算,也可查泊松分布表
因此在i代时,至少产生一个与原物种有n个基因差异的新物种个体的概率就是:
P=∑C(k,2^i)*(P2^k)*[(1-P1)^(2^i-k)] (式中k=1,2,3,……,2^i) ――式4-1
同理可用泊松定理化简为:
P=∑{(λ^k)*[e^(-λ)]/k!} (式中k=1,2,3,……,2^i;λ=(2^i)*P2)――式4-2
公式看多了头疼,我们代入些数值,举个例子看看。原公式中的M、C、R数值我们不变,生物基因数量差别很大,低等单细胞生物一般为数百个至数千个(最少的好象是300多个),我们保守一些,取m=100个。则由式2-2,单一个体子代的基因中,有一个发生变异的概率P1=0.001*0.01*0.1*100=0.0001
我们就设繁殖了i=80代(对于细菌来说,可能就是几天的事),基因差别仍然设为n=10(变异率为10%)。则由式3-2,单一个体产生满足少n个基因差异的概率可求得,其中n=10,λ=80*0.0001=0.008,求得P2=2.9375e-28
则由式4-2,所有后代种群中至少有一个新物种个体的概率可计算出,其中 λ=2^80*2.9375e-28=3.5512e-4,得到P=0.03506%。远没有原文中说的那么吓人。如果设繁殖代数i=90,可算得P=69.27%;i=100时,P=99.99……%(1500多位小数后才有变化)。计算步骤就不详细列出来了,有兴趣的朋友可以自己去算一下。这样可以看出,随分裂的不断进行,产生新物种是必然的。
当然,有朋友会说了,“你刚才的后代数量是按倍增计算的,实际上不会增长这么快,而且增长到一定程度就会停止”。是的,这的确是应该考虑的因素。考虑到这个因素,式4-1、4-2就不适用了。可以这样修正,用一个细菌形成的菌落中细菌的个数(设为j)作为参数进行计算,变为
P=∑C(k,j)*(P2^k)*[(1-P1)^(j-k)] (式中k=1,2,3,……,j) ――式4-3
P=∑{(λ^k)*[e^(-λ)]/k!} (式中k=1,2,3,……,2^i;λ=j*P2)――式4-4
这其中蕴涵着这样的假设:菌落大小不变且能长时间存在。还是用刚才的例子计算,算得的概率就小多了。但是刚才我们最多就算了100代,现在就不能只算这么少了。设i=10000,因为对于细菌等微生物来说,繁殖是很快的,按一小时分裂一次,一万小时也不过一年多点。且其菌落中个体数量是非常大的,一般每毫升中是上亿数量级的,因此我们保守设为1*10^8,即j=1e+8。
则由式3-2,λ=i*P1=10000*0.0001=1,n=10,得P2=1.1143e-7
则由式4-4,λ=j*P2=(1e+8)*(1.1143e-7)=11.143,得P=99.9986%。就是说在一年零两个月不到的时间内,该菌落中出现至少一个新物种个体的概率为99.9986%。
可看出结论是相同的,和我们的实践经验也相符合。实际科研工作中也发现,细菌等微生物的变异是很快的,新种不断在产生。sars病毒就是新种,由原来的无害且不感染人体的冠状病毒变异而来。其它的例子如细菌抗药性的产生等,都证明了这一点。
对于有性生殖,情况要复杂的多,涉及到两性之间交换遗传物质,但这种遗传物质的交换更容易使不同个体间的基因变异组合起来。这对于形成新物种更为有利。具体的计算太繁琐,计算量太大(基因多,繁殖模式也多),考虑的因素太多,通式很难列,可能需要计算机建模辅助计算,就不在这详细分析了。结论和无性生殖一致,新物种的产生绝不是偶然的极小概率事件。
文章开始处引用的这个公式,通过上面的分析试算可看出其完全是错误的,结论更是荒谬无稽。该公式出处不知道是哪里,从“以科学方法辩论”的文章中看,可能是《达尔文的黑匣子》。早就听说这是本有名的伪科学书,现在看来果不其然,居然拿这种看似科学实则荒谬的公式混淆视听,说伪科学是客气的,说造假愚弄读者也不过分。
我的计算应该说很粗糙,不排除可能有错误或者考虑不周的情况,欢迎大家批评指正。但我相信结论如果作为定性分析的依据,还是正确可靠的。
附:钱 锟:《生物学历史上最大的骗局》
英国伦敦有一位医生李察逊(Richardson),他也是胚胎学家,花了一生的时间研究人的胚胎,但他从来没有见过人胚胎有“鱼”的阶段!所以他立意要更正百多年来的错误。但是他很聪明,知道从海克尔传下来的这种“伪科学”,不是他一个人的力量可以推翻的。
??所以,他组织了十七个单位的科学家,研究了50种不同脊椎动物的胚胎及其生长过程,并且仔细观察、记录。除了海克尔用的鱼、蝾螈、龟、鸡、人、及三种哺乳动物,共四个纲的动物之外,他们还研究了圆口纲的七鳃鳗、软骨鱼中的电鳐、两栖纲的树蛙、爬行纲的鳖,哺乳动物中又加了澳大利亚的有袋类和胎盘类猫科的代表等等。
??李察逊等人终於联名在1997年8月的Anatomy & Embryology学报上发表了他们惊人的结果(同注1)。以下做简单的综述:
一、海克尔声称的“第一期”胚胎,其实并非真正的最早期胚胎。因为各纲动物从受精卵开始分裂的过程和原肠胚的形成完全不同,原肠胚以後外形上才有一些相似的地方,到後期又大不相同。所以,他说动物发育越是早期越相似,乃不尽不实之言。
二、各类动物在海克尔的第一期、也就是所谓“鱼”的时期,还是各不相同。图二是李察逊3等重新绘制的八类代表的胚胎。这些最新的资料显示出,这些动物的胚胎有相当的差异。为甚麽海克尔的原图中的八类胚胎如此相似呢?当然是动了手脚。
三、海克尔有意选择了较相近的胚胎。他选水生的蝾螈而不用青蛙代表两栖纲,是因为蝾螈本身就更像鱼。相比之下,青蛙不甚像鱼,圆口纲及软骨鱼的代表与海克尔的期望相差更远。所以不选用。
四、那麽,海克尔到底动了什麽样的手脚,以便将人的胚胎画得像鱼一样呢?原来他将人胚胎的鼻子、心脏、肝脏等大部份的内脏,及手、脚的胚芽都挖掉,再加长脊椎成尾巴!经过如此删改,想要它像甚麽都可以了(图三)。
五、根据李察逊等研究的结果,海克尔删改的不只这些,他还随意加添。例如鸡的胚胎,在这时期的眼与其他动物不同。它是没有色素的,而海克尔则将它涂黑,使它与其他动物看齐。还有,海克尔在大小比例上也随意更改,他的伸缩性可达十倍,以增加不同胚胎的相似性。
六、李察逊等人的文章还指出,海克尔刻意选用不同动物作为代表,却隐瞒这些代表的种名,使人以为同纲的动物一定都是一样的。其实不然,即使是很接近的种,它的胚胎也很可能有很大的差异。例如,不同种的鱼,它们胚胎的形态和发育的途径都可以各异。李察逊就此得出惊人结论,即“海克尔的胚胎”是生物学上最“著名”的骗局。
?? 是再度发现的骗局
??《科学》周刊的标题更指出,这个大骗局并不是首次发现,而是“再度发现”(rediscovered)。原来当年海克尔还在德国Jena大学任教期间,他伪造的这些假图就已经被人揭发。
??李察逊为了证实这是遮掩了一百多年的骗局,亲自到Jena大学去查史料。不出所料,海克尔当年被同事指控,他不但承认伪造,并且被判有罪(同注3)。所以,至今在德国的课本中找不到海克尔的图画。
??但是为甚麽在英文和中文的课本中,这些伪作流传了126年,甚至直至今天呢?李察逊说: “这才是最大的谜。”(注三)
本文作者现居旧金山,为著名美籍华裔生物学教授,曾任旧金山大学生物系主任多年,现为该系教授。
??附注:
注一:用动物模式做实验是进入临床试验前必须的步骤。各类动物之间有一定的异同,用比较方法研究更能深入了解生物之理,这是大家都知道的常识,并不需要推到4.2亿年前的假想祖先。
相反地,笔者十年前凭著蚯蚓与人类都是由一位造物主所造的信心,首创用蚯蚓作重金属污染的实验。其结果与用老鼠作的实验可以比对,与在人体的观察也非常相似。现在蚯蚓的模式已渐渐受环保及毒理学人重视。所以,在科学探讨中,动物“同源的假设”(不是已证的事实),完全可用“同一设计”的理论模式代替。换句话说,进化与创造的模式在实验科学上同样可用,但进化论理论本身却是哲学上的推论。生物学不需进化论为主导。
注二:所有脊椎动物的胚胎不是都有鳃,而是都有咽囊(phyraygeal pouches)。鱼类的咽囊发育为鳃,但在其他动物中咽囊发育成与鳃无关的结构,如内耳,和副甲状腺等。人类有尾的传说,有一部份由海克尔伪造的图而来。从图三清楚可见海克尔将脚的胚芽画成尾芽,不少人就为自己想像出尾巴来。
注三:笔者1999年暑期在中国曾接受北京某出版社副编的专访,其中问及人和鱼的胚胎比较。我建议在中学课本中删除海克尔的图画。据说这项更正在两年内可能完成。若果真如此,中国在教材上的改进要跑在英、美的前面。
引用文献:
(注1)Richardson, M. K., Hanken, J., Gooneratne, M. L., Pieau, C., Raynaud, A., Selwood, L. & Wright, G. M., 1997
There is no highly conserved embryonic stage in the vertebrates: Implications for current theories of evolution and development.
Anatomy & Embryology 196: 91-106.
(注2)Elizabeth Pennise, Sept. 5, 1997
"Haeckel's Embryos: Fraud rediscovered"
Science 277(5331): 1435.
(注3)In Brief, Sept. 6, 1997 Embryonic Fraud Lives On
New Scientist 155(2097): 23.
(注4)Jonathan Wells, 1999 Haeckel's Embryos & Evolution
The American Biology Teacher 61(5): 345-349.
(注5)Larry Witham, Jan. 25-31, 1999
Darwinism icons disputed
The Washington Times (National Weekly Edition) p.28.
(注6)Tom Abate, Feb. 7, 1999
Window on Life
San Francisco Chronicles, p.6.
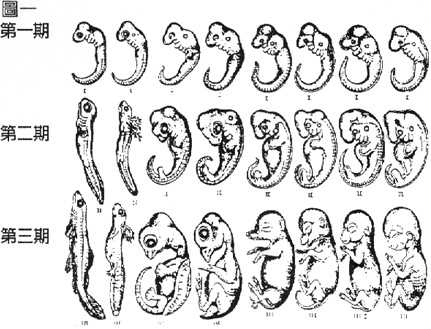
图一:海克尔手绘的胚胎比较图。从正左到右为鱼,蝾螈,龟,鸡,猪,牛,兔,及人胚胎的第一至三期。第一期也就是所谓的鱼时期,所有种类的胚胎都画成完全一样没有分别(identical)。
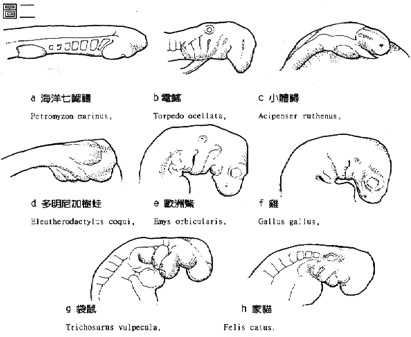
图二:李察逊等画的八种动物代表,相当於海克尔“鱼”时期的右侧图(d例外,为背侧图)。 a)海洋七鳃鳗Petromyzon marinus, b)电鳐Torpedo ocellata, c)小体鲟Acipenser ruthenus, d)多明尼加树蛙Eleutherodactylus coqui, e)欧洲鳖 Emys orbicularis, f)鸡Gallus gallus, g)袋鼠Trichosurus vulpecula, h)家猫Felis catus.
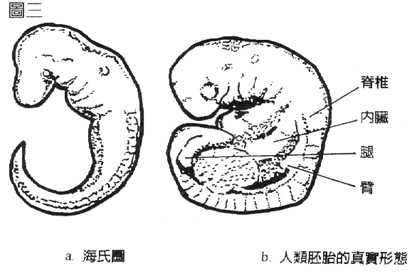
图三:海克尔删改人类胚胎的例证。左图为海克尔画的人胚胎在“鱼”的时期,右图为人胚胎真实的形态。海克尔几乎将人所有的内脏和手、脚的胚芽都挖掉。
《生物学历史上最大的骗局》摘自《海外校园》第四十期
发表评论